
N6-甲基腺苷(m6A)修饰与东说念主类疾病息息关系,它通过影响包括细胞周期、细胞行运决定和细胞稳态等1人命进度的多个方面来出动东说念主类疾病。m6A修饰的修复由Mettl3-Mettl14甲基出动酶复合物(MTC)催化2。通过Yamanaka因子(Oct4、Sox2、Klf4和c-Myc,称为OSKM)将体细胞重编程为教授性多颖悟细胞(iPSCs)3,为究诘细胞行运改动的分子机制提供了一个很好的究诘系统。之前究诘标明Mettl3的过量达不错加多m6A的水平,促进体细胞重编程为多颖悟细胞4,但Mettl14是否影响重编程以及关系作用机制尚不了了。
2022年8月9日,同济大学人命科学与本事学院高绍荣团队在Stem Cell Reports杂志上在线发表题为Mettl14-driven senescence-associated secretory phenotype (SASP) facilitates somatic cell reprogramming的究诘论文。该究诘发现,看成甲基出动酶复合物组分的Mettl14以不依赖于m6A的神情促进重编程。欺骗高通量测序本事,发现Mettl14在重编程的后期彰着上调了虚弱关系分泌表型(SASP)基因的抒发水平。这些SASP因子由未生效重编程的细胞分泌,并以旁分泌的神情促进其周围的细胞生效重编程成为iPSCs(图1)。
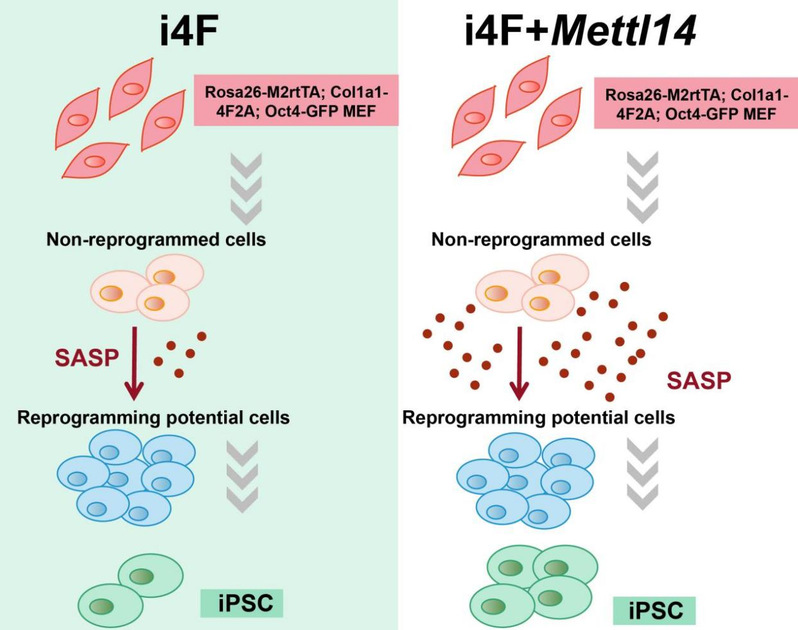
图1 Mettl14调控SASP基因
为了商酌Mettl14对体细胞重编程的影响,究诘者构建了过抒发Mettl3、过抒发Mettl14过头无活性体式的突变体质粒5(图2)。将其别离转染进重编程体系后发现,过抒发Mettl3、Mettl14以及无活性体式的Mettl14突变体均不错显赫进步重编程成果,辅导Mettl14对重编程成果的调控不依赖于m6A修饰。
究诘者在重编程经过的不同时候点,别离集聚了对照组和过抒发Mettl14 的样蓝本究诘Mettl14促进重编程的机制,并进行了转录组测序(RNA-seq)。狂妄标明,在重编程第15天,过抒发Mettl14上调的各异基因主要富集在免疫应付和细胞因子受体互相作用的基因簇。而之前的究诘标明,在虚弱细胞中,Mettl14以不依赖m6A修饰的神情调控SASP基因抒发水平6。因此,究诘者推断,在重编程晚期,Mettl14上调的这些基因为SASP基因。进一步究诘发现,在重编程晚期,这些SASP基因的抒发水平显赫升高,关联词,在建系的iPSC中果真不抒发。
为了进一步究诘SASP与重编程的关系,究诘者进行了细胞上清共培养施行及SASP因子共培养施行(图2)。狂妄标明,重编程经过的细胞上清或SASP因子(如IL6)不错促进重编程后期的细胞改动为多颖悟细胞。同期,对细胞进行碱性磷酸酶AP染色和β-Gal虚弱染色后,发现多能性细胞与虚弱细胞比例成正比,而加入虚弱胁制剂后GFP阳性细胞与虚弱细胞比例同期减少,标明重编程经过发生的虚弱对重编程经过终点进击。通过对重编程后期的细胞进行流式分选并检测其中SASP基因抒发水平发现,SASP基因在Oct4-GFP阴性细胞(未生效重编程的细胞)中的抒发水平彰着高于Oct4-GFP阳性细胞(生效重编程的细胞),究诘者推断未生效滚动为多颖悟细胞的的细胞群分泌了SASP因子,促进其周围细胞完成从体细胞向多颖悟细胞的改动。
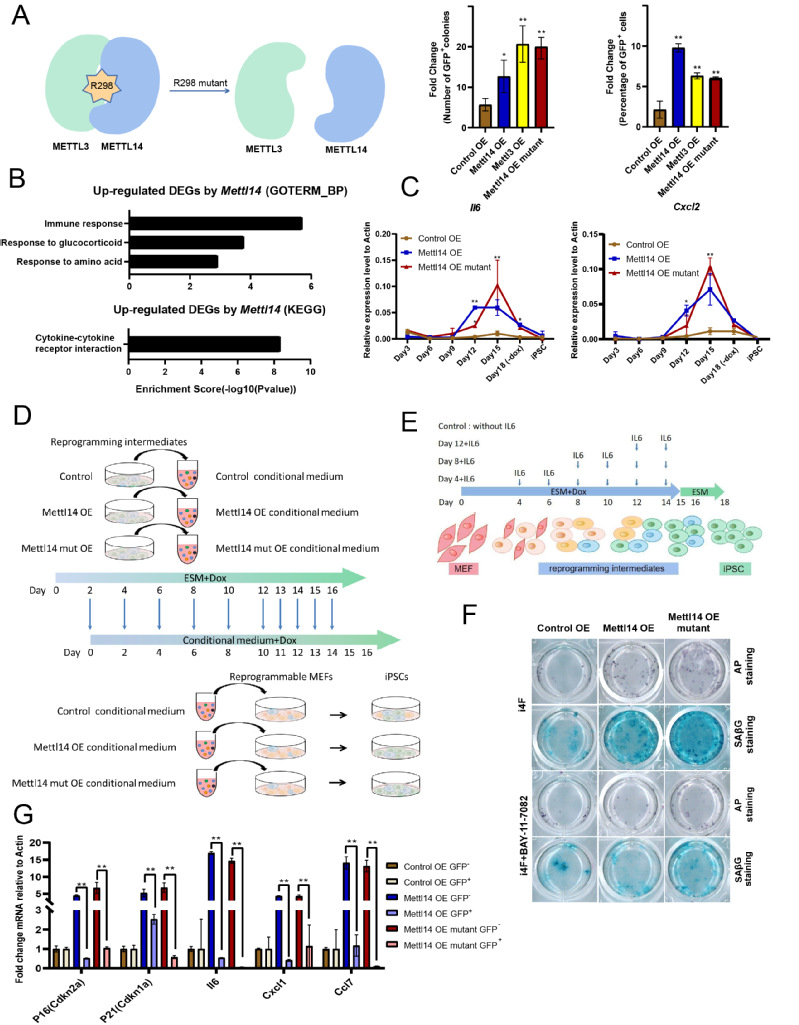
图2 体细胞重编程与细胞虚弱的关系
要而论之,该究诘揭示了Mettl3-Mettl14甲基出动酶复合物中,Mettl14对体细胞重编程的影响,考据了Mettl14在体细胞重编程经过中区别于Mettl3的作用机制,发现Mettl14在重编程教授后期调控SASP基因抒发水平,从而影响体细胞重编程的成果,同期,考据了体细胞重编程与细胞虚弱的正关系关系。该究诘为甲基出动酶寂然于m6A修饰的功能及虚弱关系分泌表型(SASP)和体外重编程之间的互相作用提供了新的见地。
本文的第一作家包括袭宸祥,孙嘉彤。高绍荣熏陶,吴丽副究诘员,郝鲁江副熏陶是本文的共同通信作家。该究诘获取了国度当然科学基金委等项打算救助。
参考文件:
1 Batista, P.J., Molinie, B., Wang, J., Qu,bt工厂网址 K., Zhang, J., Li, L., Bouley, D.M., Lujan, E., Haddad, B., Daneshvar, K., et al. (2014). m(6)A RNA modification controls cell fate transition in mammalian embryonic stem cells. Cell Stem Cell 15, 707-719.
2 Wang, X., Feng, J., Xue, Y., Guan, Z., Zhang, D., Liu, Z., Gong, Z., Wang, Q., Huang, J., Tang, C., et al. (2016). Structural basis of N(6)-adenosine methylation by the METTL3-METTL14 complex. Nature 534, 575-578.
3 Takahashi, K., and Yamanaka, S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676.
4 Chen, T., Hao, Y.J., Zhang, Y., Li, M.M., Wang, M., Han, W., Wu, Y., Lv, Y., Hao, J., Wang, L., et al. (2015). m(6)A RNA methylation is regulated by microRNAs and promotes reprogramming to pluripotency. Cell stem cell 16, 289-301.
5 Wang, X., Feng, J., Xue, Y., Guan, Z., Zhang, D., Liu, Z., Gong, Z., Wang, Q., Huang, J., Tang, C., et al. (2016). Structural basis of N(6)-adenosine methylation by the METTL3-METTL14 complex. Nature 534, 575-578.
6 Liu, P., Li, F., Lin, J., Fukumoto, T., Nacarelli, T., Hao, X., Kossenkov, A.V., Simon, M.C., and Zhang, R. (2021). m(6)A-independent genome-wide METTL3 and METTL14 redistribution drives the senescence-associated secretory phenotype. Nat Cell Biol 23, 355-365.
